Uptake, Not Excretion: What Hair Tissue Mineral Analysis Actually Measures
The most common myth in hair mineral analysis is exactly backward, and cell biology explains why.
Thank you for reading this post, don't forget to subscribe!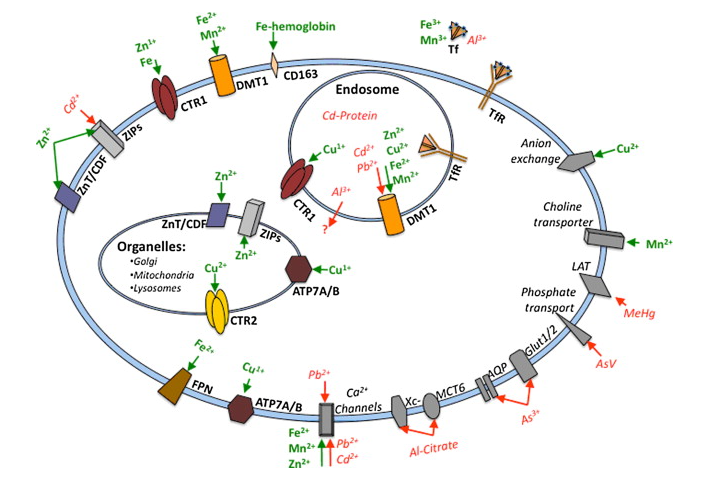
The claim that won’t die
The myth usually arrives in plain language, something close to this:
“Hair only shows what the body excretes, so if a patient excretes poorly, it won’t even show that well.”
It sounds careful. It sounds like physiology. But ask the question directly: is hair an excretory organ? Obviously not. So why is the claim repeated so confidently, often by practitioners who use hair analysis and value it? The answer is a single word, “excretion,” quietly doing two different jobs.
One word, two sciences
In toxicokinetics, “excretion” is an accounting term. Any route by which a substance leaves the body’s mass balance counts as elimination: urine, faeces, sweat, skin, and yes, hair, because once a metal is locked into a shaft that is cut or shed, that quantity has left the ledger. Toxicologists therefore list hair among elimination routes, and they are correct to.
But “excretion” carries a second, everyday meaning: active waste disposal, in which the body identifies a toxin and purposefully clears it. When the toxicology word is read through the physiology meaning, “hair reflects excretion” quietly becomes “hair reflects what you successfully detoxed,” and the false corollary drops out: poor detoxifiers won’t show it.
The error is this. The process that puts a metal into hair is not disposal. It is uptake. The hair matrix does not decide to excrete anything; it imports whatever is bioavailable in the capillary blood at the dermal papilla, using the same nutrient transporters it uses to pull in the iron, zinc and calcium it needs to build keratin. Hair records what the body took up, not what it threw away.
The follicle is a sink, not a drain
This becomes obvious once you look at what kind of tissue the hair follicle is.
The anagen hair bulb is among the most metabolically demanding structures in the body. Its matrix keratinocytes are, in the literal title of one study, a “maximally proliferative human epithelial tissue” (Purba et al. 2017), with one of the highest rates of cell division of any mammalian tissue and correspondingly high protein synthesis to build the keratin fibre (Figlak et al. 2021). Roughly 100,000 scalp follicles spin out about 12 km of shaft a year (Westgate et al. 2025). To fuel this, the follicle runs a distinctive, cancer-like metabolism (aerobic glycolysis plus glutaminolysis, with an internal Cori cycle shuttling lactate between compartments; Figlak et al. 2021), and the bulb is densely packed with active mitochondria, imaged as a circumferential “ring of fire” at the sites of fibre formation (Lemasters et al. 2017). Follicular keratinocytes carry among the highest mitochondrial concentrations of any epithelium (Wikramanayake et al. 2022; Dong et al. 2025).
None of this describes a waste outlet. It describes a voracious importer. And the demand is specifically a demand for metals: mitochondria are iron-and-manganese-loaded organelles (iron-sulphur clusters, haem cytochromes, manganese superoxide dismutase), so a maximally proliferative, mitochondria-dense tissue has a continuous hunger for iron, manganese, zinc and calcium, imported through the transferrin receptor, DMT1, the ZIP family and calcium channels. The follicle incorporates nutrients from the blood and, simultaneously, “other substances from the blood” (Florou et al. 2025), and it concentrates them, holding trace elements at multiples far above the homeostatically buffered bloodstream. A tissue that bioaccumulates minerals to that degree is, by definition, taking them up avidly. That is the opposite of excretion.
Molecular mimicry: the nutrient doors
Here is the mechanism that closes the argument. Toxic metals do not enter hair, or any tissue, through a dedicated “toxin” route. They enter through the nutrient doors, because they molecularly or ionically mimic essential elements (Bridges & Zalups 2005; Martinez-Finley et al. 2012). The follicle throws those doors open to feed its build rate, and the impostors walk through beside the real cargo.
| Transporter/receptor | Native substrate | Mimicking toxic metal |
|---|---|---|
| Transferrin receptor (TfR) | Iron, Fe³⁺ | Aluminium, manganese |
| DMT1 (divalent transporter) | Iron, Fe²⁺ | Cadmium, lead, nickel, cobalt |
| Calcium channels | Calcium, Ca²⁺ | Lead, strontium, barium |
| ZIP8 / ZIP14 (zinc transporter) | Zinc, Zn²⁺ | Cadmium |
| LAT1 / System L (amino-acid transporter) | Methionine | Methylmercury (as cysteine conjugate) |
| K⁺ channels / Na⁺,K⁺-ATPase | Potassium, K⁺ | Thallium |
| Aquaglyceroporins (AQP7 / AQP9) | Glycerol/water | Arsenic, antimony |
Two details make the point land. First, lead appears on two rows and cadmium on two: the promiscuous metals impersonate several nutrients at once, which is exactly why a normal serum iron, calcium or zinc can sit beside silent accumulation, the mimic having ridden in on the carrier the labs assumed was clean. Second, the transporters are not follicle-specific. The transferrin receptor, DMT1, calcium channels and the ZIP family sit on essentially every perfused cell, so metal in hair means that metal was in systemic circulation and being imported into cells throughout the body. Hair is a keratin tape-recording of the bioavailable metal milieu every tissue was bathed in during the same growth window.
The myth runs backward
Now return to the myth. A genuine “poor excreter” is someone who retains metal. Retention raises the circulating, bioavailable burden over time, which means more metal presented to the follicle’s transporters, not less. The myth does not just misuse a word; it points the arrow the wrong way. The patient it claims hair will miss is the patient whose hair has the most to show.
A fair word on limits
No biomarker reads whole-body or organ-specific burden, and hair is no exception. But neither is anything else. Blood is a single buffered instant of one compartment. A DEXA scan measures the spine and hip and extrapolates a T-score; it does not scan every bone. Urine, by contrast, is an actual excretion sample: it shows what the kidney has actively eliminated since the previous void. Each is a validated proxy for its own question, interpreted against reference ranges. Hair’s question is systemic bioavailable exposure and uptake integrated over roughly three months (3cm sample), and for chronic metal accumulation that is the question blood answers worst, being homeostatically clamped and reflecting only recent exposure. In a paired blood-and-hair study of healthy adults, 22 of 264 mineral measurements were normal in blood yet elevated in hair, imbalances a blood reading alone would have classified as normal (Shahverdian et al. 2026).
The one honest boundary is the deep cortical-bone vault: metal sequestered there, with a residence time of decades, is not freely in circulation, so hair, like blood, under-reads lifetime total burden. But bone is living tissue under constant remodelling. It continuously drips stored metal back into the bloodstream, surging whenever resorption accelerates, with most of the body’s lead burden held in bone against a blood half-life of about a month (Jan et al. 2015). The store that hair under-reads is the very store that keeps topping up the pool hair samples. Hair tracks the active, bioavailable burden, which is usually the clinically actionable one.
The bottom line
Hair is not a record of what the body excreted. It is a record of what the cells took up, month after month, through the very transporters that carry iron, zinc and calcium into every tissue. The myth says the worse you clear metal, the less your hair can show it. The biology says the opposite: the worse you clear it, the more you retain, the higher your bioavailable burden, and the more your hair has to report. Hair tissue mineral analysis integrates that uptake over months.
References
- Bakulski, KM, Seo, YA, Hickman, RC, Brandt, D, Vadari, HS, Hu, H & Park, SK 2020, ‘Heavy metals exposure and Alzheimer’s disease and related dementias’, Journal of Alzheimer’s Disease, vol. 76, no. 4, pp. 1215-1242, doi:10.3233/JAD-200282.
- Bridges, CC & Zalups, RK 2005, ‘Molecular and ionic mimicry and the transport of toxic metals’, Toxicology and Applied Pharmacology, vol. 204, no. 3, pp. 274-308, doi:10.1016/j.taap.2004.09.007.
- Dong, TR, Li, YJ, Jin, SY, Yang, FL, Xiong, RX, Dai, YQ, Song, XZ & Guan, CP 2025, ‘Progress on mitochondria and hair follicle development in androgenetic alopecia: relationships and therapeutic perspectives’, Stem Cell Research & Therapy, vol. 16, article no. 44, doi:10.1186/s13287-025-04182-z.
- Figlak, K, Williams, G, Bertolini, M, Paus, R & Philpott, MP 2021, ‘Human hair follicles operate an internal Cori cycle and modulate their growth via glycogen phosphorylase’, Scientific Reports, vol. 11, article no. 20761, doi:10.1038/s41598-021-99652-8.
- Florou, VA, Manaprasertsak, A, Slyusarenko, M, Amend, SR, Kazi, JU, Hammarlund, EU & Pienta, KJ 2025, ‘Human hair as a diagnostic tool in medicine’, Biochemistry and Biophysics Reports, vol. 43, article no. 102129, doi:10.1016/j.bbrep.2025.102129.
- Jan, AT, Azam, M, Siddiqui, K, Ali, A, Choi, I & Haq, QMR 2015, ‘Heavy metals and human health: mechanistic insight into toxicity and counter defense system of antioxidants’, International Journal of Molecular Sciences, vol. 16, no. 12, pp. 29592-29630, doi:10.3390/ijms161226183.
- Lemasters, JJ, Ramshesh, VK, Lovelace, GL, Lim, J, Wright, GD, Harland, D & Dawson, TL 2017, ‘Compartmentation of mitochondrial and oxidative metabolism in growing hair follicles: a ring of fire’, Journal of Investigative Dermatology, vol. 137, no. 7, pp. 1434-1444, doi:10.1016/j.jid.2017.02.983.
- Liu, Z, Shen, J, Carbrey, JM, Mukhopadhyay, R, Agre, P & Rosen, BP 2002, ‘Arsenite transport by mammalian aquaglyceroporins AQP7 and AQP9’, Proceedings of the National Academy of Sciences, vol. 99, no. 9, pp. 6053-6058, doi:10.1073/pnas.092131899.
- Martinez-Finley, EJ, Chakraborty, S, Fretham, SJB & Aschner, M 2012, ‘Cellular transport and homeostasis of essential and nonessential metals’, Metallomics, vol. 4, no. 7, pp. 593-605, doi:10.1039/c2mt00185c.
- Purba, TS, Brunken, L, Peake, M, Shahmalak, A, Chaves, A, Poblet, E, et al. 2017, ‘Characterisation of cell cycle arrest and terminal differentiation in a maximally proliferative human epithelial tissue: lessons from the human hair follicle matrix’, European Journal of Cell Biology, vol. 96, no. 7, pp. 632-641, doi:10.1016/j.ejcb.2017.03.011.
- Shahverdian, A, Torabzadeh, S, Watts, D, Porwollik, S, Padilla, M, Lee, JS, Perez, G & Jafari, M 2026, ‘Comparison of mineral levels in blood and hair samples of healthy adults: evaluating the clinical utility of hair mineral analysis’, Biological Trace Element Research, vol. 204, no. 3, pp. 1915-1927, doi:10.1007/s12011-025-04793-w.
- Tallkvist, J, Bowlus, CL & Lönnerdal, B 2003, ‘Effect of iron treatment on nickel absorption and gene expression of the divalent metal transporter (DMT1) by human intestinal Caco-2 cells’, Pharmacology & Toxicology, vol. 92, no. 3, pp. 121-124, doi:10.1034/j.1600-0773.2003.920303.x.
- Westgate, GE, Grohmann, D & Sáez Moya, M 2025, ‘Hair longevity: evidence for a multifactorial holistic approach to managing hair aging changes’, Journal of Clinical Medicine, vol. 14, no. 6, article no. 1894, doi:10.3390/jcm14061894.
- Wikramanayake, TC, Chéret, J, Sevilla, A, Birch-Machin, M & Paus, R 2022, ‘Targeting mitochondria in dermatological therapy: beyond oxidative damage and skin aging’, Expert Opinion on Therapeutic Targets, vol. 26, no. 3, pp. 233-259, doi:10.1080/14728222.2022.2049756.
No Comments
Sorry, the comment form is closed at this time.